- 1 Sur le nom du site, lire Maureille et al. (2001).
1Situé sur la commune de Montignac (Dordogne), le gisement moustérien de Regourdou1 est localisé sur la rive gauche de la Vézère, à 205 m d’altitude, au sommet de la colline qui abrite la grotte de Lascaux (Piveteau 1959). Il a été découvert et partiellement « exploité » par Roger Constant son propriétaire (Piveteau 1963) puis scientifiquement fouillé de 1960 à 1964 par Eugène Bonifay (Bonifay et al. 2007). La datation des principaux faciès sédimentaires est incertaine. D’après l’analyse des associations fauniques (Delpech 1996), la couche 4, qui livre les restes humains, serait estimée à environ 75 ka (soit la seconde moitié de OIS 5). Certains auteurs (Vallois 1965 ; Vandermeersch et Trinkaus 1995) l’associent plutôt à la fin du Würm I ou au début de l’interstade Würm I/II (soit le début de l’OIS 3) d’après l’analyse sédimentologique ou microfaunique (Simard 1968). L’un des principaux intérêts du site réside dans le fait qu’il livre un squelette néandertalien bien conservé (Regourdou 1) qui pourrait être la conséquence d’une des plus vieilles sépultures d’Europe (Maureille et Vandermeersch 2007). Cette dernière fait encore l’objet de controverses, notamment en ce qui concerne certains gestes funéraires des Néandertaliens comme celui du prélèvement intentionnel de la boîte crânienne (Bonifay 2002 ; Maureille 2004 ifay et al. 2007 versus Madelaine et al.)
2À la fin des fouilles (1964) les membres supérieurs et le tronc sont majoritairement représentés chez Regourdou 1. De la ceinture pelvienne et des membres inférieurs ne sont alors connus qu’un sacrum, deux petits fragments d’ischium et de pubis droits, une patella gauche et 18 os des pieds. Mais, heureusement, une révision-inventaire des collections fauniques du site, commencée en 2008 par Stéphane Madelaine, a permis la découverte de nouveaux restes humains, dont un important fragment de diaphyse fémorale à droite, la patella droite, un fragment distal de tibia gauche et les deux tiers de la fibula à gauche et surtout des fragments importants d’iliums et d’ischiums droits et gauches (Madelaine et al. 2008).
3Avec l’association de ces nouveaux restes de bassin au sacrum, à l’ischium et au pubis connus, la ceinture pelvienne de Regourdou compte désormais parmi les plus complètes de ce taxon fossile avec Kebara 2 (Bar-Yosef et Vandermeersch 1991). Sa description permet donc de mieux cerner la variabilité morphologique intra-populationnelle de cette région anatomique particulièrement fragile. De plus, ce bassin nous autorisera dans le futur à discuter certaines implications morpho-fonctionnelles pouvant être déduites des caractéristiques anatomiques de ces fossiles.
4Dans la nuit du 22 au 23 septembre 1957, des « invités » de R. Constant mettent au jour des ossements humains (Piveteau 1959 ; Bonifay et Vandermeersch 1962 ; Bonifay et al. 2007) appartenant à un individu : Regourdou 1. Un calcaneus plus volumineux que celui connu à droite chez Regourdou 1 sera ultérieurement mis au jour dans le gisement. Il est logiquement attribué à un second individu (Regourdou 2) par Vandermeersch et Trinkaus (1995). L’industrie lithique, réalisée sur des matières premières locales ou provenant des alluvions de la Vézère, présente dans la couche stratigraphique livrant Regourdou 1 est issue d’un système technique discoïde axé sur la production de pointes pseudo-levallois (Turq, com. pers.) et non pas de supports de type Quina (Bonifay 1965).
5Après leur découverte, les ossements humains ont été malheureusement souvent déplacés (du Muséum National d’Histoire Naturelle au site de Regourdou puis au Musée de l’Homme ou au Laboratoire d’Anthropologie de l’Université Bordeaux 1) avant d’être acquis par le Musée d’Art et d’Archéologie du Périgord en 1984 (Madelaine et al. 2008). Ils ont été parfois manipulés sans précaution ce qui explique, par exemple, l’état de conservation actuel des premières vertèbres cervicales par rapport à ce qui a été décrit par Piveteau (1964, 1966). Dans le cadre de travaux de récolement des collections des musées de France, le MNP inventorie la totalité du matériel présent dans ses collections. Lors des travaux sur les collections fauniques du site de Regourdou, l’un d’entre nous (S. M.) découvre, le 21 mai 2008, une diaphyse fémorale droite appartenant à un individu néandertalien. D’autres découvertes (S. M., Bruno Maureille et Nadia Cavanhié) permettront de reconstituer une grande partie du squelette appendiculaire inférieur de Regourdou 1 (Madelaine et al. 2008).
6L’âge au décès de Regourdou 1 a été estimé par Vandermeersch et Trinkaus (1995), à l’aide des stades d’attrition dentaire de Molnar (1971). L’éruption et les degrés d’usure ont ainsi permis aux auteurs d’estimer que Regourdou 1 est un jeune adulte qui n’aurait pas plus de 25 ans.
7En le comparant à l’ensemble des Néandertaliens européens dont le sexe a pu être établi (Trinkaus 1980), Vandermeersch et Trinkaus (ibidem) soulignent que Regourdou 1 présente une mosaïque de caractères métriques, parfois dans la variabilité néandertalienne « féminine » (le diamètre de la tête radiale), parfois dans la variabilité néandertalienne « masculine » (longueurs humérales, talaires, radiales) et parfois entre les deux sexes (taille générale, longueur de l’extrémité distale de l’humérus, morphologie de la branche supérieure du pubis). Pour cette raison, les auteurs concluent que « its sex is best regarded as indeterminate » (Vandermeersch et Trinkaus 1995 - p. 441). D’autres auteurs considèrent Regourdou 1 en tant qu’individu masculin. C’est le cas de Vallois (1965) qui écrit que l’étude des os a montré qu’ « il s’agissait là d’un sujet sans doute masculin » ou de Churchill et Formicola (1997) qui inscrivent Regourdou dans leur étude en tant que masculin. Gomez-Olivencia et collaborateurs (2007) ont, quant à eux, déterminé Regourdou comme masculin à partir de fonctions discriminantes établies par Wescott (2000) et basées sur l’axis, au sein de populations modernes. L’utilisation d’analyses mises en place sur des populations actuelles pour déterminer le sexe de populations néandertaliennes est connu comme un biais important (Calgagno 1981). Cela empêche donc de considérer ce dernier résultat comme probant. Pour le moment, nous préférons nous en tenir à l’hypothèse de Vandermeersch et Trinkaus (1995) et considérer que nous ne connaissons pas le sexe de ce fossile.
8Enfin, l’étude des membres supérieurs a permis à Trinkaus et Vandermeersch (1995 - p. 472) d’estimer, selon Trotter et Gleser (1952), la stature de Regourdou 1 entre 164,6 et 167,5 cm.
- 2 L’ensemble de la description anatomique a comme référence l’anatomie selon Kamina (Kamina 1995 et 1 (...)
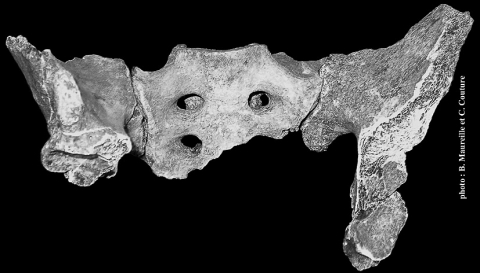
9Parmi les nouvelles découvertes de 2008, plusieurs fragments d’ilium (os ilium) et d’ischium (os ischii) ont attiré l’attention. Les deux fragments d’ilium présentant leur surface auriculaire, cela a permis de les associer avec certitude au sacrum de Regourdou 1 (fig. 1). Les fragments d’ischium, par leur ressemblance avec le fragment ischiatique connu sur Regourdou 1, peuvent aussi être rapportés à ce même individu. Finalement, les pièces identifiées lors du récolement permettent donc de reconstituer la ceinture pelvienne de Regourdou 1 : le sacrum associé aux iliums et à des fragments d’ischium et de pubis (os pubis).
Figure 1 - Le sacrum de Regourdou 1 en connexion anatomique avec les iliums droit et gauche.
Figure 1 - The sacrum of Regourdou 1, associated with the two ilia.
- 3 L’ischium droit et le sacrum ont été décrits par François Marchal (1997) mais n’ont pas été publiés (...)
10(fig. 2)
11Seuls les deux premières vertèbres sacrées et l’aileron droit de la troisième sont conservés : en effet, l’os est brisé suivant une ligne oblique qui, antérieurement, va du bord supérieur du deuxième trou sacré gauche à l’extrémité inférieure de la surface auriculaire droite. Le fragment est particulièrement bien conservé et ne présente aucune fracture ou déformation. L’os présente une usure marquée ainsi qu’une « patine » particulière qu’on retrouve sur l’ischium droit, lui aussi présent avant 2008. Cette « patine » pourrait être due à l’application d’un produit consolidant après la fouille à un traitement post-fouille particulier (D. Armand, com. pers.) ou à une opération de moulage mal contrôlée.
Figure 2 - Le sacrum de Regourdou 1.
Figure 2 - The sacrum of Regourdou 1.
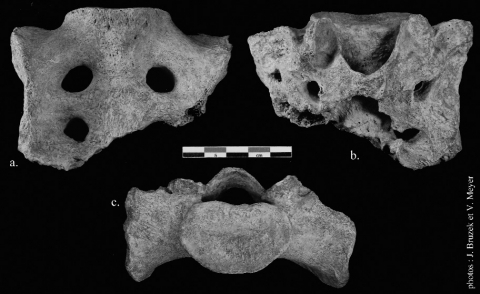
a. Face antérieure ; b. Face postérieure ; c. Base.
a. Anterior view ; b. Posterior view ; c. Basal view.
12(fig. 2a)
13Le sacrum est hypobasal. On remarque que les ailerons sacrés (alae ossis sacri) sont peu étalés latéralement ce qui induit une étroitesse de l’os (104 mm de large) qui reste néanmoins dans la variabilité moderne (Been et al. 2010). Par ailleurs, l’aileron gauche est nettement plus projeté (latéralement et antéropostérieurement) que le droit (la projection latérale de l’aileron gauche est de 55 mm contre 49 mm à droite). La base du sacrum (basis ossis sacri) et le promontoire (promontorium) montrent une morphologie moderne. Les crêtes transversales (crista sacralis lateralis) sont peu visibles car très usées. La première vertèbre est plus épaisse que la deuxième. Les premiers foramens sacrés (foramina sacralia pelvina) sont plus importants que l’unique deuxième foramen conservé. Notons aussi que la ligne de suture (linea transversae) entre la première et la deuxième vertèbre est encore visible. Cela ne permet pas d’indiquer l’âge au décès de l’individu, en effet, la persistance d’une ligne de suture est fréquente chez les adultes (Williams et al. 1995).
14(fig. 2b)
15Médialement, l’orifice supérieur du canal sacré (canalis sacralis) est marqué et profondément ouvert vers le bas. Le premier tubercule spinal est absent en raison de l’érosion générale de l’os. La crête sacrée médiane (crista sacralis mediana) est brisée en dessous de ce tubercule.
16Les processus articulaires (processus articularis superior) de la première vertèbre sont déformés et présentent un allongement antéro-postérieur. Le processus droit est cassé, ce qui peut expliquer l’impression de non individualisation qui ressort d’une première observation. Le tubercule transversal droit est assez développé. Mais il est impossible de le comparer au gauche qui est érodé.
17Les tubercules sacrés postérieurs (tuberositas sacralis) sont trop érodés pour être analysés. Les premiers foramens sacrés postérieurs (foramina sacralia dorsalia) sont seuls visibles et ne présentent aucune caractéristique particulière.
18Seule la surface articulaire droite (facies auricularis) est complète (à gauche, la moitié inférieure est manquante). Les deux surfaces sont réniformes, à hile plutôt dorsal, bien que la morphologie de la surface droite soit plus sinueuse. La forte érosion de cette zone empêche une description plus précise. On observe toutefois un sillon péri-auriculaire sur les deux surfaces. Placé antéro-inférieurement, il est particulièrement marqué sur la surface latérale gauche.
19(fig. 2c)
20La base ne présente aucun caractère particulier autre que l’asymétrie entre les ailerons droit et gauche déjà soulignée (cf. supra). Néanmoins, en regardant le sacrum en vue supérieure, on observe, à nouveau, le développement important du canal sacré.
21Le sacrum ne présente aucun caractère permettant de le différencier d’un sacrum moderne. Cela va dans le sens des résultats déjà obtenus par Trinkaus (1984) qui souligne qu’il y a peu de différence entre un sacrum néandertalien et moderne. Seule l’ouverture du canal sacré est notable mais il faut la rapporter à la variabilité de la morphologie moderne du sacrum (Pirro et al. 2007). Peut-être faut-il corréler cette ouverture profonde du canal sacré avec le hiatus sacré fortement ouvert chez certains Néandertaliens, notamment Shanidar 1 et La-Chapelle-aux-Saints (Trinkaus 1983) ? Finalement, il semble y avoir une grande variabilité de taille au sein des sacrums néandertaliens (tabl. 1). En effet, si Regourdou 1 présente un sacrum peu puissant, Heim (1982), dans sa description de La Ferrassie 1, a mis en avant la robustesse de cet os, le qualifiant de très épais tandis que dans leur étude des restes des fossiles de Spy, Fraipont et Lohest (1887) le décrivent comme étroit. Le sacrum de Kebara 2, bien que plus large et plus long, ne diffère pas d’un sacrum moderne (Rak 1991 - p. 151). Enfin, décrivant les sacrums de Shanidar 1, 3 et 4, Trinkaus (1983) note que la morphologie de ces derniers ne s’éloigne pas de celle des autres Néandertaliens ni d’une architecture moderne.
Tableau 1 - Variabilité métrique des sacrums néandertaliens.
Table 1 - Metrical variability of neandertal sacra.

22(fig. 3a, b et e ; fig. 4a et b ; fig. 4f et g)
23L’os coxal droit de Regourdou 1 est représenté par un grand fragment d’ilium, l’ischium et un fragment de pubis.
24(fig. 3a, b et e)
25La partie centrale de l’ilium droit est présente, l’aile (ala ossis ilii) et la crête iliaque (crista iliaca) ne sont pas préservées. Bien qu’on ne puisse donc conclure sur la taille du coxal droit, de nombreux caractères de robustesse générale peuvent être décrits, au niveau des insertions musculaires et de l’épaisseur de l’os (voir plus loin). Il a été retrouvé sous la forme de six fragments qui ont été assemblés. L’os n’est pas fossilisé. S’il est bien conservé on observe aussi de nombreuses cassures fraîches. La structure interne de l’os est intacte et pourra se prêter à des études plus poussées [un travail sur l’organisation des structures trabéculaires apporterait ainsi beaucoup d’informations biomécaniques sur le pelvis (Volpato 2007)]. L’absence de l’aile a aussi pour conséquence qu’il est impossible de la comparer avec le corps de l’ilium (corpus osis ilii). Rak a bien décrit la morphologie particulière de l’ilium de Kebara 2, comme une morphologie en champignon (« mushroom-like appearance » Rak 1991 - p. 147) ; un déséquilibre de taille entre le corps et l’aile de l’ilium étant à l’origine de cette morphologie particulière (Rak et Arensburg 1987 ; Rak 1991). Dans le cas de Regourdou 1, il est impossible de rapprocher la morphologie de l’ilium d’une morphologie en champignon, comme celle de Kebara 2, ou encore de celle d’Amud 1 (Endo et Kimura 1970), dont la taille de l’aile est moins disproportionnée par rapport à la taille générale de l’ilium, ou de celle de La Ferrassie 1 dont la position externe de l’aile est frappante selon Heim (1982).
Figure 3 - Les iliums de Regourdou 1.
Figure 3 - The ilia of Regourdou 1.
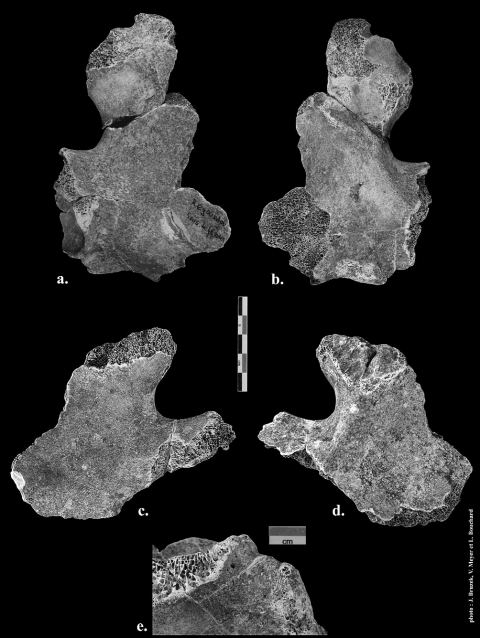
a. Ilium droit, face externe ; b. Ilium droit, face interne ; c. Ilium gauche, face externe ; d. Ilium gauche, face interne ; e. L’épine iliaque antéro-inférieure, fortement déviée médialement, vue interne.
a. Right, external view ; b. Right, internal view ; c. Left, external view ; d. Left, internal view ; e. Antero-inferior iliac spine and its medial deviation, internal view.
26(fig. 3a et e)
27Toute la partie inférieure de l’aile est conservée : la partie supérieure de l’acétabulum (acetabulum), la partie supérieure de la grande incisure ischiatique (incisura ischiadica major) et la zone postérieure de la surface auriculaire (facies auricularis) tout comme l’épine iliaque antéro-inférieure (spina iliaca anterior inferior) sont présentes.
28Au niveau de l’acétabulum, seule la partie iliaque de la surface semi-lunaire (facies lunata), adjacente à l’épine iliaque antéro-inférieure, est préservée. Elle est lisse, légèrement courbée et mesure environ 42,5 mm de long sur 34 mm de large. On n’observe ni la limite avec l’arrière-fond, ni le bord supérieur de la surface semi-lunaire. La partie supérieure de l’acétabulum présente une gouttière bien marquée. Cette gouttière correspond à la zone d’insertion du chef réfléchi du muscle droit fémoral. L’insertion de ce muscle est donc très profonde, ce qui pourrait indiquer une musculature développée.
29Cette hypothèse est confortée par la morphologie de l’épine iliaque antéro-inférieure (fig. 4e). En effet, cette dernière est remarquablement puissante (sa longueur est de 29 mm ; son épaisseur est de 12 mm) et proéminente comparée à un bassin moderne. Or, elle correspond à l’insertion du chef direct du muscle droit fémoral, ce qui implique donc ici aussi une musculature plus puissante que chez l’Homme anatomiquement moderne. Par ailleurs, cette épine est orientée vers l’intérieur de la cavité pelvienne. Cette déviation médiale de l’épine se retrouve chez d’autres spécimens néandertaliens comme par exemple Kebara 2 (Rak 1991 ; Rak et Arensburg 1987 ; Marchal 1997). La profondeur inter-épine, qui peut permettre d’évaluer la puissance de l’épine antéro-inférieure n’est pas mesurable, en l’absence de l’épine antéro-supérieure (spina iliaca anterior superior). Cette profondeur est évaluée à 16,5 mm chez Kebara 2 (Rak 1991). D’après la synthèse de la morphologie pelvienne des Néandertaliens selon Marchal (1997), l’épine iliaque antéro-inférieure est bien séparée de l’acétabulum, massive et présente une déviation marquée, particulièrement chez La-Chapelle-aux-Saints, Kebara 2, Feldhofer 1 et Tabun C1. Amud 1 présente une épine iliaque dont la massivité dépasse celle des autres Néandertaliens (Endo et Kimura 1970). Les auteurs soulignent la déviation médiale et notent qu’elle n’est pratiquement pas observée chez l’Homme moderne (Endo et Kimura 1970 - p. 301).
30L’espace entre l’acétabulum et l’épine iliaque antéro-inférieure présente une fosse au niveau de l’insertion du ligament ilio-fémoral. Au dessus de l’acétabulum, la ligne glutéale inférieure (linea glutealis inferior) est très rugueuse. Cela pourrait indiquer la présence d’un muscle petit fessier puissant.
31Le pilier iliaque n’est pas observable, en raison de la cassure de l’os en face antérieure qui empêche d’évaluer l’épaisseur de l’aile. La position et l’épaisseur de ce pilier ont été discutées par Endo et Kimura (1970), par McCown et Keith (1939) ou par Vandermeersch (1981). Notons que Trinkaus, en 1984, observe que ce pilier est plus faiblement développé chez l’Homme de Néandertal que chez l’Homme anatomiquement moderne. C’est aussi la conclusion de Marchal (1997 - p. 100) qui note que le pilier iliaque des Néandertaliens est « moyennement développé, en position antérieure et oblique sur l’aile ». Cette observation rejoint l’étude de Vandermeersch (1981 - p. 236) sur les hommes de Qafzeh : le pilier iliaque de Qafzeh 9 est en arrière, tandis que le pilier sur le bassin néandertalien est en position plus avancée.
32(fig. 3b)
33Comme pour la face externe, seule la partie inférieure de l’ilium est représentée, avec la ligne arquée (linea arcuata), la surface auriculaire et l’épine iliaque antéro-inférieure. Aucune partie de la crête iliaque n’a pu être observée.
34En raison de l’absence de l’aile, il est difficile de discuter la profondeur de la fosse iliaque (fossa iliaca). Celle-ci est très variable chez l’Homme de Néandertal. Marchal (1997) note que cette profondeur va en s’intensifiant au cours de l’évolution. Il parle de fosse iliaque « moyennement concave à très concave » chez les Néandertaliens et souligne la concavité très marquée de la fosse iliaque chez l’Homme anatomiquement moderne, particulièrement chez les sujets masculins (Marchal 1997 - p. 105).
- 4 Les données métriques modernes ont été établies à titre indicatif à partir d’un échantillon de 30 i (...)
35La ligne arquée est bien délimitée, ce qui provoque une angulation importante entre la fosse et la surface quadrilatère. La frontière entre le petit et le grand bassin est donc bien marquée. L’ilium y est particulièrement puissant, caractère déjà observé chez Kebara 2 (Rak 1991) et La Ferrassie 1 (Heim 1982). On a pu en mesurer l’épaisseur au niveau de l’incisure ischiatique : environ 35 mm chez Regourdou 1 contre 25 mm pour Kebara 2 (Rak 1991) et 22,4 mm en moyenne chez l’Homme anatomiquement moderne [17,67 ; 28,75]4.
36L’insertion du muscle iliaque est bien creusée. Il s’insère aussi sur l’épine iliaque antéro-inférieure qui, comme précisé précédemment, est très robuste. Il semblerait donc que les muscles de la région ilio-fémorale soient puissants.
37Sur la face interne de l’épine iliaque antéro-inférieure, une petite fosse est observable. Ceci est à mettre en relation avec l’ensemble de cette région anatomique, caractérisée par des zones d’insertions musculaires marquées.
38La région pré-auriculaire est bien conservée, sauf au niveau de sa partie médio-dorsale où la corticale est « dénudée ». Il n’y a aucune trace de sillon pré-auriculaire ni de sillon para-glénoïdaire. L’épine iliaque postéro-inférieure (spina iliaca posterior inferior) n’est pas observable.
39L’os est fragmenté juste entre la région pré-auriculaire et la surface auriculaire. Le bras antérieur de cette dernière est détérioré contrairement au bras postérieur dont la largeur a pu être mesurée (31-32 mm). La longueur de la surface a pu être évaluée à 54 mm. La surface est bien lisse et caractéristique d’un individu jeune. L’érosion de la surface sacro-pelvienne de l’ilium empêche d’estimer l’âge au décès de l’individu suivant les méthodes établies par Schmitt et Broqua (2000), Buckberry et Chamberlain (2002) ou Schmitt (2005) . Il est difficile d’évaluer la position de la surface articulaire, en raison de l’absence de la crête et de l’état général de la région auriculaire (facies auricularis). Cette position est variable chez l’Homme de Néandertal. Elle est haute chez Kebara 2 ou Feldhofer 1 (Marchal 1997), basse chez La-Chapelle-aux-Saints (Boule 1912).
- 5 Ces deux mesures ont été prises sur les moulages du Laboratoire d’Anthropologie des Populations Pas (...)
40Deux mesures (Gaillard 1960) ont pu être prises sur cette zone : la longueur spino-sciatique (69,6 mm) et la longueur spino-auriculaire (71 mm). La puissance de l’os au niveau de la surface auriculaire est de 25 mm (25 mm chez Kebara 2, 28 mm chez Feldhofer 1, 20,39 en moyenne chez l’Homme anatomiquement moderne [10,75 ; 27,02]5.
41La grande incisure ischiatique est incomplète. Il est donc difficile de statuer sur son étroitesse ou sa profondeur. Néanmoins, le bras postérieur et la portion de bras antérieur présents permettent d’observer que l’incisure est plutôt étroite. D’après Endo et Kimura (1970), l’incisure ischiatique d’Amud 1 est remarquablement large mais normalement profonde, proche de la morphologie de Feldhofer 1. Chez Tabun, elle est large et peu profonde (McCown et Keith 1939). Enfin, chez La Ferrassie 1, Heim (1982) souligne surtout que le bord antérieur de la grande incisure ischiatique est rectiligne. Cela n’a pas pu être observé chez Regourdou 1, le bras (ou bord) antérieur étant absent.
42(fig. 4a et b)
43Le dernier fragment associé au coxal droit de Regourdou 1 est un ischium presque complet : on observe la partie inférieure de l’acétabulum, le corps de l’ischium (corpus ossis ischii) et la tubérosité ischiatique (tuber ischiadicum). Il manque la partie ichiatique de la branche ischio-pubienne (ramus ossis ischii) et l’épine sciatique (spina ischiadica) est érodée. L’os est remarquablement robuste. La cassure de ce fragment au niveau de l’acétabulum pourrait correspondre à la zone d’ossification, peut-être encore fragile à l’âge de l’individu ; par ailleurs, la tubérosité est complète. On peut donc considérer que la longueur du fragment, soit 80 mm, correspond à la longueur de l’ischium anatomique (Schultz 1930). L’os présente une usure remarquable, qui peut être due à un traitement post-fouille particulier. Ainsi, on observe de nombreuses traces qui pourraient être la conséquence de moulages (E. Pubert com. pers.).
Figure 4 - Les ischiums et le pubis de Regourdou 1.
Figure 4 - The ischia and the pubis of Regourdou 1.

a. Ischium droit, face externe ; b. ischium droit, face interne ; c. et d. Ischium gauche, fragment d’acétabulum ; e. Ischium gauche, tubérosité ischiatique ; f et g : Pubis droit, fragment de branche supérieure.
a. Right ischium, external view ; b. Right ischium, internal view ; c. and d. Left ischium, acétabulum fragment ; e. Left, ischiatic tuberosity ; f. and g. : Right pubis, fragment of the superior ramus.
44(fig. 4a)
45La tubérosité ischiatique est massive et mesure 58 mm de long. En vue postéro-interne, elle présente une dépression marquée qui correspond à l’insertion du muscle biceps fémoral. Cette dépression est à mettre en relation avec la puissante rugosité en position supéro-externe, où s’insère le muscle semi-membraneux. Rappelons que dans sa description de La Ferrassie 1, Heim (1982) remarque la puissance de cette insertion. Bien que moins marquées, les insertions des muscles semi-tendineux et grand adducteur sont fortes. La crête transverse est peu marquée et se présente sous la forme d’une concavité mousse, tandis que la crête longitudinale est très marquée. Cette morphologie est déjà décrite chez Kebara 2 par Rak (1991). En ce qui concerne la position et la morphologie du sillon du muscle obturateur interne, Trinkaus (1996) a souligné la variabilité importante de cette région morphologique chez les Néandertaliens et les Hommes anatomiquement modernes. Dans le cas présent, le sillon est en position craniale (ou postéro-supérieure), passant entre la tubérosité et l’épine ischiatique. La tubérosité est située plus latéralement sur l’ischion que ce qu’on observe chez l’homme moderne ; néanmoins, sa latéralisation est moins frappante que chez Kebara (Rak 1991). Supérieurement, l’insertion du muscle carré fémoral est marquée par une protubérance dans la continuité de la tubérosité.
46La partie postérieure de l’acétabulum est préservée. La partie postéro-inférieure de la surface semi-lunaire (dont la largeur ne dépasse pas 22 mm) est saillante et bien en relief sur la partie supérieure à la tubérosité. F. Marchal (com. pers.) la compare à une corne et souligne que : « sa saillie se projette presque au niveau du cadre du foramen obturé ». La gouttière sous-acétabulaire est étroite et profonde. Si cette étroitesse est observée chez les Homo erectus et de nombreux hommes actuels (Marchal 1997), la profondeur est une caractéristique commune à Regourdou 1, La Ferrassie 1 (Heim 1982) et La-Chapelle-aux-Saints (Boule 1912). Cela doit être lié avec l’insertion du muscle obturateur externe. En son milieu, on observe une dépression plus ou moins circulaire, qui pourrait être, d’après F. Marchal (1997), la zone d’insertion de la racine postérieure du ligament rond. La surface rétro-acétabulaire est plane, plus étroite que chez l’Homme moderne - mesure n° M14.1- (Bräuer 1988). Son épaisseur est de 34 mm contre une moyenne moderne de 36,53 mm [29,29 ; 43,67]6. Elle est estimée à 33 mm chez Kebara 2 par Rak (1991). Cette étroitesse serait apparemment une caractéristique des pelvis des représentants fossiles du genre Homo (Marchal 1997, 2000). Toutefois, le caractère « plat » de cette zone semble spécifique aux Néandertaliens (Marchal 1997). On observe de nombreux foramens nourriciers (une dizaine environ) au niveau de la zone tubéro-acétabulaire.
47(fig. 4b)
48L’épine ischiatique est cassée mais on peut observer néanmoins que sa base est puissante. La surface quadrilatère est vaste et légèrement concave. La tubérosité produit un bourrelet osseux conséquent qui correspond ici à l’insertion du ligament sacro-tubéral.
49(fig. 4f et g)
- 6 Bien que l’on ne soit pas certain de la localisation exacte du fragment de pubis, la section en a é (...)
50Parmi les pièces isolées, on trouve un fragment de branche supérieure du pubis. Il serait peut-être à localiser du côté droit en raison de la position du pecten (pecten ossis pubis) et de la forme de la section de l’os. Ce fragment est assez massif et montre un pecten bien marqué qui souligne une insertion puissante du ligament pectiné. Le pubis néandertalien est caractérisé par une élongation de la branche supérieure marquée par rapport aux hommes modernes (Stewart 1960 ; Trinkaus 1976, 1982, 1984 ; Rak 1990, 1991 ; Marchal 2000), élongation estimée à 20 % par Marchal (2000), mais aussi par une section particulière : la branche supérieure du pubis présente en effet un important aplatissement cranio-caudal (Trinkaus 1976 ; Rak 1990). La section du fragment de branche supérieure de Regourdou a pu être décrite6. On retrouve la morphologie typique des pubis néandertaliens : un amincissement supéro-inférieur (fig. 5). Notons la similitude entre la section transverse du pubis de Kebara 2, Shanidar 1 et celle de Regourdou 1 : on retrouve bien chez ce dernier la structure en « T couché » décrite sur les premiers par Marchal (1997 - p. 142 ; 2000).
Figure 5 - Comparaison des sections transverses de la branche supérieure du pubis des Néandertaliens et des Hommes anatomiquement modernes.
Figure 5 - Comparison of the transverse sections of the superior pubic ramus of Neandertals and modern Humans. After Marchal 2000.

D’après Marchal 2000.
51(fig. 3c et d ; fig. 4c, d et e)
52L’os coxal gauche de Regourdou 1 est représenté par un fragment d’ilium, associé à deux fragments d’ischium.
53(fig 3c et d)
54L’ilium gauche est très bien représenté. Il est reconstitué à partir de deux fragments et est mieux conservé que le droit. Par ailleurs, l’ensemble de l’os a le même aspect très puissant qu’à droite. Toute la partie inférieure de l’aile est conservée. Le fond de la grande incisure ischiatique est intact, seules les extrémités des bras supérieurs et inférieurs sont endommagées. Toutefois, la cavité acétabulaire est plus fragmentée qu’à droite. La structure interne de l’os est bien conservée et fera l’objet d’une étude ultérieure. De nombreuses cassures fraîches sont aussi notées.
55(fig. 3c)
56La ligne glutéale inférieure est marquée, on y retrouve les rugosités indiquant une insertion plus puissante du muscle petit fessier. La grande incisure ischiatique est ici bien observable. Elle est relativement fermée et symétrique. La bordure antérieure de la surface auriculaire et le bras antérieur de la grande incisure forment un arc composé de Genovés (Genovés 1959).
57Peu de caractères particuliers sont observés sur cette face qui présente les mêmes caractéristiques générales qu’à droite.
58(fig. 3d)
59L’ilium est présent de la surface auriculaire jusqu’à l’épine iliaque antéro-inférieure qui n’est pas conservée.
60La profondeur de la fosse iliaque est peu marquée. La ligne arquée est très prononcée, on retrouve l’angulation importante entre le plan de la fosse iliaque et celui de la surface quadrilatère.
61La surface auriculaire est très bien conservée. Elle est très large et présente une surface irrégulière. On y observe une fente en son milieu. L’origine de cette fente est douteuse, elle pourrait être d’origine taphonomique ou liée à l’ossification incomplète du cartilage de la surface auriculaire (Scheuer et Black 2000). Si tel est le cas, cela permettrait de confirmer que Regourdou 1 est un jeune adulte. Une dépression bien marquée est présente sur la surface pré-auriculaire. On peut la rapporter à l’insertion du ligament sacro-iliaque ventral.
62(fig. 4c, d et e)
63L’ischium est représenté par deux fragments isolés. Un fragment d’acétabulum (fig. 4c et d) et un fragment de tubérosité ischiatique (fig. 4e).
64Seule la partie inférieure de la tubérosité est conservée. Massive, elle présente les mêmes caractéristiques qu’à droite mais une usure importante empêche de décrire toutes les insertions musculaires aussi précisément. L’insertion du semi-membraneux est bien marquée. Aucune crête, longitudinale ou transverse, n’est préservée. Le bourrelet de la tubérosité correspondant à l’insertion du ligament sacro-tubéral est présent ; la zone rétro-tubérositaire est caractérisée à ce niveau par une dépression ponctuée de petits foramens. L’insertion du carré-fémoral est marquée mais plus saillante qu’à droite. Toutefois, elle est incomplète, il faut donc considérer cette observation avec précaution.
65Le fragment d’acétabulum isolé est associé à l’ischium gauche ; inférieurement, on y note une douzaine de foramens nourriciers. La corne postérieure de l’acétabulum est très proéminente, comme cela a déjà été constaté sur l’ischium droit. La gouttière sous-acétabulaire est marquée mais il n’y a pas de dépression comme cela a été noté à droite. De nombreux foramens nourriciers sont présents au niveau de la cavité acétabulaire ainsi que de petites marques rouges, probablement d’origine taphonomique (D. Armand, com. pers.)
66Complété par les nouvelles pièces découvertes, le bassin de Regourdou 1 peut aujourd’hui être décrit et intégré à la variabilité morphologique néandertalienne. Il est caractérisé par une puissance marquée, notamment au niveau de l’ilium et de la tubérosité ischiatique. Bien qu’elle ait déjà été notée chez La-Chapelle-aux-Saints (Boule 1912) et La Ferrassie 1 (Heim 1982), il faut souligner que cela ne semble pas être commun à tous les Néandertaliens. En effet, Trinkaus (1982, 1983) décrit les restes pelviens de Shanidar 1, 3 et 4 comme larges mais peu robustes. Cette puissance s’observe non seulement sur les insertions musculaires, qui peuvent témoigner d’une musculature développée, mais aussi sur l’épaisseur de l’os (par exemple, au niveau de l’incisure ischiatique) et de la massivité de certaines structures anatomiques (comme l’épine iliaque antéro-inférieure). On a pu observer que cette morphologie n’est pas généralisée à l’ensemble de la ceinture pelvienne. En effet, le sacrum ne montre aucun trait morphologique associable à une massivité particulière. Par ailleurs, il ne présente pas de caractéristiques permettant de le distinguer d’un sacrum moderne.
67On remarque aussi sur ces os coxaux de nombreux caractères déjà décrits sur les bassins néandertaliens connus à ce jour. La-Chapelle-aux-Saints, La Ferrassie 1, Kebara 2, Feldhofer 1, Krapina 207 et 209, Tabun C1, Amud 1 ou Shanidar 1, 3 et 4 (pour ne citer que les plus complets) ont fait l’objet de nombreuses publications qui permettent de connaître la variabilité pelvienne chez l’Homme de Néandertal. Notre description permet d’ajouter Regourdou 1 à cet inventaire, ce qui présente un intérêt indéniable. En effet, cela permet de constater la récurrence de certains caractères. La déviation médiale de l’épine iliaque antéro-inférieure (peut-être associée à une incisure inter-épineuse antérieure profonde, non préservée chez Regourdou 1), l’étroitesse de la surface rétro-iliaque ou la profondeur de la gouttière sous-acétabulaire, entre autres, ont été fréquemment décrites et se retrouvent sur le bassin de Regourdou 1. En ce qui concerne l’incisure inter-épineuse antérieure profonde, Majo (2000) a observé que ce caractère était présent chez les individus immatures de deux sites moustériens : La Ferrassie et Djebel Qafzeh. Cet auteur souligne que, dans les deux cas, cette échancrure épineuse est profonde chez les individus immatures (LF 6 et LF 8 ; Q10 et Q21) mais que seuls les Néandertaliens présentent ce caractère à l’âge adulte. Elle s’oppose ainsi à la description de Qafzeh 9 par Vandermeersch (1981) qui observe, chez cet individu adulte, une échancrure profonde. Par ailleurs, ce dernier souligne que si on retrouve l’échancrure inter-épineuse profonde chez Qafzeh 9, il n’en va pas de même pour la déviation médiale de l’épine iliaque antéro-inférieure. Pour lui, ce caractère est constant chez les Néandertaliens et très rarement présent chez les Hommes anatomiquement modernes (Vandermeersch 1981 - p. 236). Néanmoins, Marchal (1997) souligne que chez certains Néandertaliens (Amud et Krapina) cette déviation rentre dans la variabilité moderne.
68Par ailleurs, malgré la faiblesse de l’échantillon se rapportant au pubis, il a été possible d’étudier la section de ce dernier (fig. 5). La branche supérieure du pubis de Regourdou présente un aplatissement cranio-caudal marqué. Cet aplatissement se retrouve chez Amud, qui est décrit comme « long et mince » (Endo et Kimura 1970), chez Kebara (Rak 1990) et Shanidar (Trinkaus 1983). En 1976, Trinkaus montre que cette morphologie est commune aux Néandertaliens, non seulement du Proche-Orient mais aussi d’Europe. Le pubis de Regourdou présente donc des traits caractéristiques à l’ensemble des Néandertaliens. Différentes hypothèses ont été proposées pour expliquer la morphologie particulière du pubis néandertalien : adaptation obstétricale (Trinkaus 1984 ; Rosenberg 1988 ; Trevathan, 1988), biomécanique (Rak et Arensburg 1987 ; Marchal 2000), adaptation au froid (Ruff 1994) ou plésiomorphie (Marchal 2000) sont les principales explications proposées. En 1989, Anderson propose un bon récapitulatif de ces différentes hypothèses.
69Finalement, l’absence de pubis complet ne permet pas d’observer un des caractères les plus marquants de l’os coxal des Néandertaliens : l’élongation du pubis. Néanmoins, certains traits morphologiques observés chez Regourdou 1 permettent de le différencier des bassins d’Hommes anatomiquement modernes.
70Ainsi, si la morphologie générale de l’ilium et de l’ischium néandertalien est semblable à une morphologie moderne, comme l’ont déjà souligné Trinkaus (1983, 1984), Vandermeersch (1981) ou McCown et Keith (1939), il existe certains traits qui peuvent différencier un coxal d’Homme moderne d’un coxal de Néandertalien (la déviation médiale de l’épine iliaque antéro-inférieure, l’étroitesse de la surface rétro iliaque ou la profondeur de la gouttière sous-acétabulaire). Compte tenu de notre effectif de comparaison (trente individus récents), il est difficile de prendre en compte la variabilité morphologique moderne. Une étude comparative plus poussée, faite à partir d’un échantillon élargi d’Hommes anatomiquement modernes, permettra de comprendre si ces caractères peuvent être considérés comme typiques des Néandertaliens. Notons que, parmi les caractères énoncés, aucun n’est aussi frappant que peuvent l’être ceux qui distinguent un pubis moderne d’un pubis néandertalien. On peut alors considérer que, par rapport au pubis, les iliums et ischiums sont proches de la morphologie moderne.
71De nombreux spécialistes ont proposé un sexe masculin pour Regourdou 1 mais il convient de rester prudent quant à l’interprétation des divers caractères pelviens observés. En effet, en l’absence de mesures classiques sur les coxaux, on ne veut pas conclure trop hâtivement sur la détermination sexuelle de Regourdou 1. Cette question sera abordée ultérieurement avec des travaux plus avancés.
72Par ailleurs, on compare ici un individu néandertalien européen, Regourdou 1, à des individus issus de différents sites d’Eurasie et particulièrement à plusieurs individus du Proche-Orient, comme Tabun C1, Kebara 2, Amud 1 ou Shanidar 1, 3 et 4. Cela doit inciter à la prudence. Si on a pu mettre en évidence une variabilité morphologique importante au sein des Néandertaliens, il faut peut-être modérer cette observation eu égard aux différents débats sur les Néandertaliens orientaux et leurs relations avec les Hommes modernes du Proche-Orient ou les Néandertaliens européens. Ainsi, diverses études ont mis ou mettent en avant d’une part la difficulté de définir deux populations au Porche-Orient et d’autre part l’absence d’homogénéité anatomique entre les Néandertaliens européens et asiatiques (par exemple McCown et Keith 1939 ; Thoma 1957, 1958, 1965 ; Vandermeersch 1989 ; Mann 1995 ; Arensburg 1991 ; Arensburg et Belfer-Cohen 1998 ; Quam et Smith 1998 ; Rak 1998 ; Vandermeersch 2007 ; Tillier et al. 2008).
- 7 Cette notion de paradoxe obstétrical (en anglais : obstetrical dilemna) est aujourd’hui nuancée par (...)
73L’étude du bassin a un intérêt tout particulier quand on s’intéresse à l’évolution, de par l’implication de la ceinture pelvienne à la fois dans la bipédie et dans la parturition. On se place ici au sein de ce que de nombreux auteurs ont appelé « le paradoxe obstétrical » (Abitbol 1996 ; Rosenberg et Trevathan 2002 ; Raynal et al. 2005) : l’adaptation à la bipédie nécessite un changement de conformation du complexe fémoro-pelvien qui s’oppose à l’adaptation morphologique du bassin à l’encéphalisation (et donc à l’accouchement de nouveau-nés dont la capacité crânienne est élevée7). L’étude de ce nouveau pelvis au sein de la variabilité néandertalienne permettra ainsi d’initier une réflexion sur la question de l’obstétrique néandertalienne, question qui a fait l’objet de nombreuses études (Trinkaus 1984 ; Rosenberg 1988 ; Anderson 1989) dont certaines, très récentes, ont donné des résultats contradictoires (Weaver et Hublin 2009 versus Ponce de Leon et al. 2008). L’existence d’un nouveau bassin néandertalien est ainsi l’occasion d’approfondir le débat sur la parturition néandertalienne.
74La ceinture pelvienne de Regourdou 1 est composée d’un sacrum (représenté par les deux premières vertèbres sacrées) qui s’articule avec les deux iliums droit et gauche (fig. 1), tous deux bien conservés mais ne présentant ni l’aile ni la crête iliaque, de l’ischium droit presque complet et très bien conservé, de deux fragments d’ischium gauche et enfin d’une partie de la branche supérieure du pubis droit. La morphologie générale des os coxaux et du sacrum ne s’éloigne pas d’une morphologie moderne. Cependant, on remarque la présence de caractères déjà décrits sur d’autres pelvis néandertaliens : la déviation de l’épine iliaque antéro-inférieure ; la profondeur de la gouttière sous-acétabulaire ; une zone rétro-acétabulaire non seulement étroite mais aussi plate ; enfin, un amincissement cranio-caudal de la branche supérieure du pubis. Ainsi, avec la découverte et la description de la ceinture pelvienne de Regourdou 1, l’effectif des pelvis néandertaliens est augmenté d’un nouveau spécimen, celui-ci comptant parmi les plus complets ; cela met d’abord en évidence l’existence d’une variabilité morphologique au sein de l’échantillon néandertalien. C’est aussi l’occasion de mettre en exergue certains caractères communs à tous ces bassins ; caractères qui pourraient, grâce à une étude plus précise, être considérés comme dérivés.
Véronique Merlin-Anglade, conservatrice du Musée d’Art et d’Archéologie de Périgueux, nous a très aimablement prêté les restes de bassin de Regourdou 1 pour que nos recherches puissent se dérouler dans des conditions favorables en fonction des contraintes doctorales à l’Université Bordeaux 1. Nous lui exprimons notre reconnaissance pour cette compréhension qui nous est indispensable. Nous remercions aussi très chaleureusement Guy Marchesseau, responsable des collections du Paléolithique au MAAP, pour son accueil, son aide et la valorisation d’un patrimoine périgordin exceptionnel. Nous sommes naturellement très reconnaissant à Jean-Jacques Cleyet-Merle, directeur du Musée national de Préhistoire (Les Eyzies-de-Tayac), qui a permis que ces nouvelles recherches sur les vestiges de Regourdou se concrétisent et se développent dans le cadre d’un doctorat (V. M.). Nous espérons qu’elles seront à la hauteur de ses espérances et au profit des deux institutions muséales de Dordogne. Merci également à l’ensemble de l’équipe du Musée national de Préhistoire pour l’accueil toujours chaleureux. Merci enfin à Laureline Bouchard pour son aide photographique, Anne-Marie Tillier, Dominique Armand, Isabelle Crevecoeur et Eric Pubert de l’UMR5199 PACEA pour nos discussions fructueuses.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}